фотосинтез
ФОТОСИНТЕЗ
образование зелеными растениями и некоторыми бактериями орг. веществ с использованием энергии солнечного света. Происходит при участии пигментов (у растений хлорофиллов). В основе Ф. лежат окислительно-восстановит. реакции, в которых электроны переносятся от донора (напр., H2O, H2S) к акцептору (CO2) с образованием восстановленных соед. (углеводов) и выделением O2 (если донор электронов H2O), S (если донор электронов, напр., H2S) и др.
Ф. — один из самых распространенных процессов на Земле, обусловливает круговорот в природе углерода, O2 и др. элементов. Он составляет материальную и энергетич. основу всего живого на планете. Ежегодно в результате Ф. в виде орг. вещества связывается ок. 8∙1010 т углерода, образуется до 1011 т целлюлозы. Благодаря Ф. растения суши образуют ок. 1,8∙1011 т сухой биомассы в год; примерно такое же количество биомассы растений образуется ежегодно в Мировом океане. Тропич. лес вносит до 29% в общую продукцию Ф. суши, а вклад лесов всех типов составляет 68%. Ф. высших растений и водорослей — единственный источник атм. O2.
Возникновение на Земле ок. 2,8 млрд. лет назад механизма окисления воды с образованием O2 представляет собой важнейшее событие в биол. эволюции, сделавшее свет Солнца главным источником-своб. энергии биосферы, а воду — практически неограниченным источником водорода для синтеза веществ в живых организмах. В результате образовалась атмосфера совр. состава, O2 стал доступным для окисления пищи (см. дыхание), а это обусловило возникновение высокоорганизов. гетеротрофных организмов (применяют в качестве источника углерода экзогенные орг. вещества).
Ок. 7% орг. продуктов Ф. человек использует в пищу, в качестве корма для животных, а также в виде топлива и строит. материала. Ископаемое топливо — тоже продукт Ф. Его потребление в кон. 20 в. примерно равно приросту биомассы.
Общее запасание энергии солнечного излучения в виде продуктов Ф. составляет ок. 1,6∙1021 кДж в год, что примерно в 10 раз превышает совр. энергетич. потребление человечества. Примерно половина энергии солнечного излучения приходится на видимую область спектра (длина волны λ от 400 до 700 нм), которая используется для Ф. (физиологически активная радиация, или ФАР). ИК излучение не пригодно для Ф. кислородвыделяющих организмов (высших растений и водорослей), но используется некоторыми фотосинтезирующи-ми бактериями.
В связи с тем, что углеводы составляют осн. массу продуктов биосинтетич. деятельности растений, хим. уравнение Ф. обычно записывают в виде:
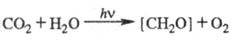
Для этой реакции  469,3 кДж/моль, понижение энтропии 30,3 Дж/(К∙моль),
469,3 кДж/моль, понижение энтропии 30,3 Дж/(К∙моль), 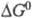 −479 кДж/моль. Квантовый расход Ф. для одноклеточных водорослей в лаб. условиях составляет 8–12 квантов на молекулу CO2. Утилизация при Ф. энергии солнечного излучения, достигающего земной поверхности, составляет не более 0,1% всей ФАР. Наиб. продуктивные растения (напр., сахарный тростник) в среднем за год усваивают ок. 2% энергии падающего излучения, а зерновые культуры — до 1%. Обычно суммарная продуктивность Ф. ограничена содержанием CO2 в атмосфере (0,03–0,04% по объему), интенсивностью света и температурой. Зрелые листья шпината в атмосфере нормального состава при 25 °C на свету насыщающей интенсивности (при солнечном освещении) дают неск. литров O2 в час на грамм хлорофилла или на килограмм сухого веса. Для водорослей Chlorella pyrenoidosa при 35 °C повышение концентрации CO2 от 0,03 до 3% позволяет повысить выход O2 в 5 раз, такая активация является предельной.
−479 кДж/моль. Квантовый расход Ф. для одноклеточных водорослей в лаб. условиях составляет 8–12 квантов на молекулу CO2. Утилизация при Ф. энергии солнечного излучения, достигающего земной поверхности, составляет не более 0,1% всей ФАР. Наиб. продуктивные растения (напр., сахарный тростник) в среднем за год усваивают ок. 2% энергии падающего излучения, а зерновые культуры — до 1%. Обычно суммарная продуктивность Ф. ограничена содержанием CO2 в атмосфере (0,03–0,04% по объему), интенсивностью света и температурой. Зрелые листья шпината в атмосфере нормального состава при 25 °C на свету насыщающей интенсивности (при солнечном освещении) дают неск. литров O2 в час на грамм хлорофилла или на килограмм сухого веса. Для водорослей Chlorella pyrenoidosa при 35 °C повышение концентрации CO2 от 0,03 до 3% позволяет повысить выход O2 в 5 раз, такая активация является предельной.
Бактериальный Ф. и общее уравнение Ф. Наряду с Ф. высших растений и водорослей, сопровождаемым выделением O2, в природе осуществляется бактериальный Ф., в котором окисляемым субстратом является не вода, а др. соединения, обладающие более выраженными восстановит. свойствами, напр. H2S, SO2. Кислород при бактериальном Ф. не выделяется, напр.:
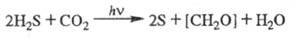
Фотосинтезирующие бактерии способны использовать не только видимое, но и ближнее ИК излучение (до 1000 нм) в соответствии со спектрами поглощения преобладающих в них пигментов — бактериохлорофиллов. Бактериальный Ф. не имеет существенного значения в глобальном запасании солнечной энергии, но важен для понимания общих механизмов Ф. Кроме того, локально бескислородный Ф. может вносить существенный вклад в суммарную продуктивность планктона. Так, в Черном море количество хлорофилла и бактериохлорофил-ла в столбе воды в ряде мест приблизительно одинаково.
Учитывая данные о Ф. высших растений, водорослей и фотосинтезирующих бактерий, обобщенное уравнение Ф. можно записать в виде:
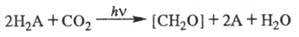
А — кислород в случае высших растений и водорослей, S либо др. элементы — в бактериальном Ф.
Мол. механизм Ф. и структура фотосинтетич. аппарата.
С использованием изотопных меток показано, что источником O2 в Ф. является только вода:

Ф. пространственно и во времени разделяется на два сравнительно обособленных процесса: световую стадию окисления воды и темновую стадию восстановления CO2 (рис. 1). Обе эти стадии осуществляются у высших растений и водорослей в специализир. органеллах клетки — хлоропластах. Исключение — синезеленые водоросли (цианобактерии), у которых нет аппарата Ф., обособленного от цитоплазматич. мембран.
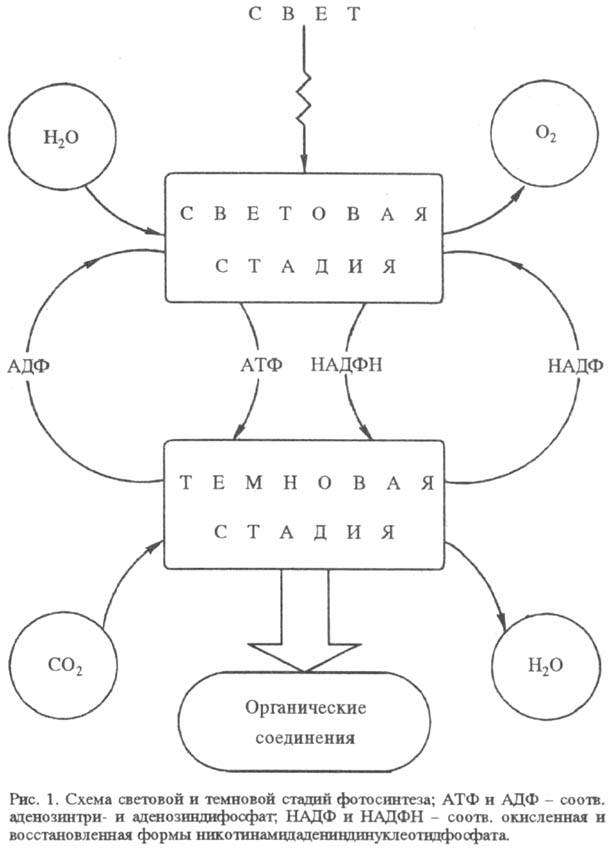
Хлоропласт, представляющий собой замкнутую структуру, отделенную от остальной части клетки оболочкой, заключает в себе весь фотосинтетич. аппарат. Световая стадия реализуется в мембранных структурах хлоропласта (т. наз. тилакои-дах), тогда как темновая стадия происходит в жидком содержимом хлоропласта (строме) при участии водорастворимых ферментов. У фотосинтезирующих бактерий хлоропласты отсутствуют, но световая стадия также осуществляется в мембранных образованиях — в т. наз. хроматофорах.
Световая стадия. Миним. функциональная единица, еще способная осуществлять световую стадию Ф.,- тилакоид. Он представляет собой микроскопич. плоский диск, образованный белковолипидными мембранами, в которых находятся пигменты. В эти мембраны встроены все компоненты, необходимые для окисления воды, восстановления кофермента никотинамиддинуклеотидфосфата (НАДФ) до НАДФН и синтеза АТФ из аденозиндифосфата. Световая стадия инициируется поглощением кванта света пигментами, организованными в спец. светособирающие комплексы. Среди пигментов преобладает хлорофилл а. К вспомогат. пигментам относятся хлорофилл b, каротиноиды и др. Наличие светособирающей структуры из неск. сотен или десятков молекул пигментов на каждый фотохимически активный (реакционный) центр на 2–3 порядка увеличивает сечение захвата излучения и обеспечивает возможность Ф. при слабом освещении.
Часть вспомогат. пигментов, спектрально наиб. близких к фотохимически активному хлорофиллу, непосредственно окружает каждый из реакционных центров, образуя т. наз. антенны.
Высокая эффективность переноса возбуждения от молекулы, поглотившей квант, к фотохим. центру определяется спектр. свойствами и структурной организацией пигментов светособирающего комплекса и антенны, окружающей фотохим. центр. Эти пигменты обеспечивают передачу возбуждения за время менее 100 пс в пределах времени жизни синглетно возбужденного состояния хлорофилла.
В реакц. центре Ф., куда почти со 100%-ной вероятностью переносится возбуждение, происходит первичная реакция между фотохимически активной молекулой хлорофилла а (у бактерий — бактериохлорофилла) и первичным акцептором электрона (ПА). Дальнейшие реакции в тилакоидных мембранах происходят между молекулами в их осн. состояниях и не требуют возбуждения светом. Эти реакции организованы в электронтранспортную цепь — последовательность фиксированных в мембране переносчиков электрона. В электронтранс-портной цепи высших растений и водорослей содержится два фотохим. центра (фотосистемы), действующих последовательно (рис. 2), в бактериальной электронтранспортной цепи — один (рис. 3).
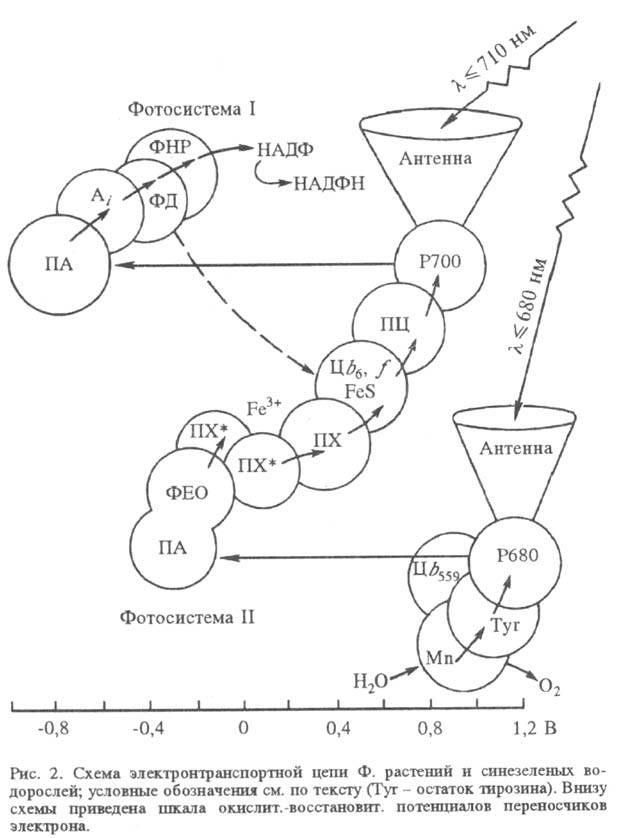
В фотосистеме II высших растений и водорослей синглетно возбужденный хлорофилл а в центре Р680 (число 680 обозначает, что максимум спектральных изменений системы при возбуждении светом находится вблизи 680 нм) отдает электрон через промежуточный акцептор к феофитину (ФЕО, безмагниевый аналог хлорофилла), образуя катион-радикал 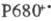 . Анион-радикал восстановленного феофитина служит далее донором электрона для связанного пластохинона (ПХ*; отличается от убихинонов заместителями в хиноидном кольце), координированного с ионом Fe3+ (в бактериях имеется аналогичный Fe3+-убихинонный комплекс). Далее электрон переносится по цепи, включающей свободный пластохинон (ПХ), присутствующий в избытке по отношению к остальным компонентам цепи, затем цитохромы (Ц) b6 и f, образующие комплекс с железо-серным центром, через медьсодержащий белок пластоцианин (ПЦ; мол. м. 10400) к реакционному центру фотосистемы I.
. Анион-радикал восстановленного феофитина служит далее донором электрона для связанного пластохинона (ПХ*; отличается от убихинонов заместителями в хиноидном кольце), координированного с ионом Fe3+ (в бактериях имеется аналогичный Fe3+-убихинонный комплекс). Далее электрон переносится по цепи, включающей свободный пластохинон (ПХ), присутствующий в избытке по отношению к остальным компонентам цепи, затем цитохромы (Ц) b6 и f, образующие комплекс с железо-серным центром, через медьсодержащий белок пластоцианин (ПЦ; мол. м. 10400) к реакционному центру фотосистемы I.
Центры 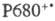 быстро восстанавливаются, принимая электрон через ряд промежут. переносчиков от воды. Образование O2 требует последоват. четырехкратного возбуждения реакционного центра фотосистемы П и катализируется мембранным комплексом, содержащим Mn.
быстро восстанавливаются, принимая электрон через ряд промежут. переносчиков от воды. Образование O2 требует последоват. четырехкратного возбуждения реакционного центра фотосистемы П и катализируется мембранным комплексом, содержащим Mn.
Хлорофилл α в фотосистеме I, имеющий максимум поглощения вблизи 700 нм (центр Р700), является первичным фотовозбуждаемым донором электрона, который он отдает первичному акцептору (ПА; его природа однозначно не установлена), а затем, через ряд промежут. переносчиков (Ai) — растворимому белку ферредоксину (ФД), восстанавливающему с помощью фермента ферредоксин-НАДФ-редуктазы (ФНР) НАДФ до НАДФН. Катион-радикал окисленного пигмента 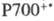 восстанавливается пластоцианином.
восстанавливается пластоцианином.
В зрелых хлоропластах имеются граны (стопки тилакоидов), в мембранах которых присутствуют все компоненты злектронтранспортной цепи, и т. наз. агранальные тилакоиды, не содержащие фотосистемы II.
Благодаря асимметрич. расположению компонентов электронтранспортной цепи относительно плоскости мембраны при разделении зарядов между хлорофиллом в каждом из двух фотосинтетич. центров и акцептором электрона на тилако-идной мембране создается разность электрич. потенциалов (плюс — на внутренней, минус — на внешней ее стороне). Перенос электрона пластохиноном сопровождается транспортом протонов, которые захватываются снаружи тилакоида при восстановлении пластохинона и освобождаются внутрь тилакоида при окислении пластогидрохинона. Перенос электронов сопряжен с синтезом АТФ из аденозиндифосфата (АДФ) и неорг. фосфата. Предполагают, что обратный транспорт протонов из тилакоидов в строму через белковый сопрягающий фактор (H+-АТФ-синтетазу) сопровождается образованием АТФ.

Фотосистема I может действовать автономно без контакта с системой II. В этом случае циклич. перенос электрона (на схеме показан пунктиром) сопровождается синтезом АТФ, а не НАДФН. Образующиеся в световой стадии кофермент
НАДФН и АТФ используются в темновой стадии Ф., в ходе которой снова образуется НАДФ и АДФ.
Электронтранспортные цепи фотосинтезирующих бактерий в основных своих чертах аналогичны отдельным фрагментам таковых в хлорогшастах высших растений. На рис. 3 показана электронтранспортная цепь пурпурных бактерий.
Темновая стадия Ф. Все фотосинтезирующие организмы, выделяющие O2, а также некоторые фотосинтезирующие бактерии сначала восстанавливают CO2 до фосфатов Сахаров в т. наз. цикле Калвина. У фотосинтезирующих бактерий встречаются, по-видимому, и др. механизмы. Большинство ферментов цикла Калвина находится в растворимом состоянии в строме хлоропластов.
Упрощенная схема цикла показана на рис. 4. Первая стадия — карбоксилирование рибулозо-1,5-дифосфата и гидролиз продукта с ооразованием двух молекул 3-фосфоглицериновой кислоты. Эта C3-кислота фосфорили-руется АТФ с образованием 3-фосфоглицероилфосфата, который затем восстанавливается НАДФН до глицеральдегид-3-фосфата. Полученный триозофосфат затем вступает в ряд реакций изомеризации, конденсации и перегруппировок, дающих 3 молекулы рибулозо-5-фосфата. Последний фосфорилируется при участии АТФ с образованием риоу-лозо-1,5-дифосфата и, т. обр., цикл замыкается. Одна из 6 образующихся молекул глицеральдегид-3-фосфата превращается в глюко-зо-6-фосфат и используется затем для синтеза крахмала либо выделяется из хлоропласта в цитоплазму. Глицеральдегид-3-фосфат может также превращаться в 3-глицеро-фосфат и затем в липиды. Триозофосфаты, поступающие из хлоропласта, превращаются в осн. в сахарозу, которая переносится из листа в др. части растения.
В одном полном обороте цикла Калвина расходуется 9 молекул АТФ и 6 молекул НАДФН для образования одной молекулы 3-фосфоглицериновой кислоты. Энергетич. эффективность цикла (отношение энергии фотонов, необходимых для фотосинтеза АТФ и НАДФН, к DG0 образования углевода из CO2) с учетом действующих в строме хлоропласта концентраций субстратов составляет 83%. В самом цикле Калвина нет фотохим. стадий, но световые стадии могут косвенно влиять на него (в т. ч. и на реакции, не требующие АТФ или НАДФН) через изменения концентраций ионов Mg2+ и H+, а также уровня восстановленности ферредоксина.
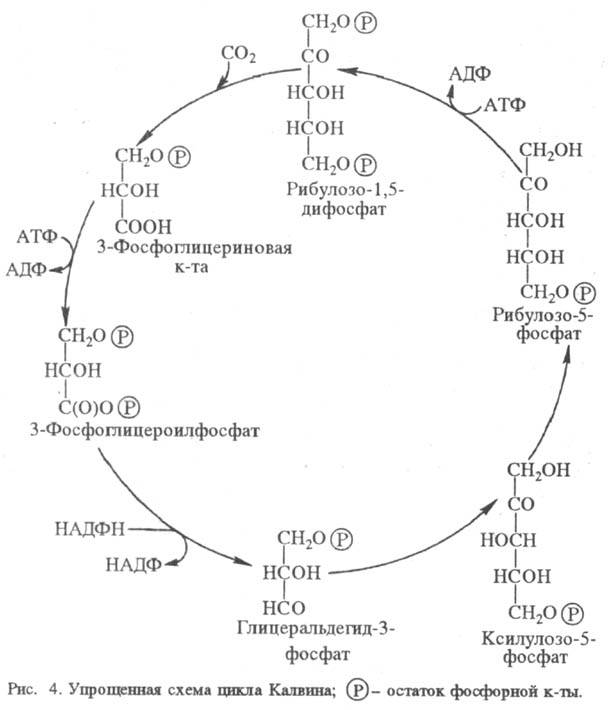
Некоторые высшие растения, приспособившиеся к высокой интенсивности света и к теплому климату (напр., сахарный тростник, кукуруза), способны предварительно фиксировать CO2 в дополнит. C4-цикле. При этом CO2 сначала включается в обмен четырехуглеродных дикарбоновых кислот, которые затем декарбоксилируются там, где локализован цикл Калвина. C4-Цикл характерен для растений с особым анатомич. строением листа и разделением функций между двумя типами клеток: мезофильных, где сосредоточено карбоксилирование фосфоенолиировиноградной кислоты, и клеток обкладки сосудистого пучка, где функционирует цикл Калвина. Образующаяся в C4-цикле щавелевоуксусная кислота восстанавливается НАДФН до яблочной, которая перемещается в клетки сосудистой обкладки и здесь подвергается окислит. декарбоксили-рованию, образуя пировиноградную кислоту, CO2 и НАДФН. Два последних используются в цикле Калвина, а пировиноградная кислота возвращается в C4-цикл (рис. 5). Физиол. смысл C4-цикла состоит в запасании CO2 и повышении, т. обр., общей эффективности процесса.
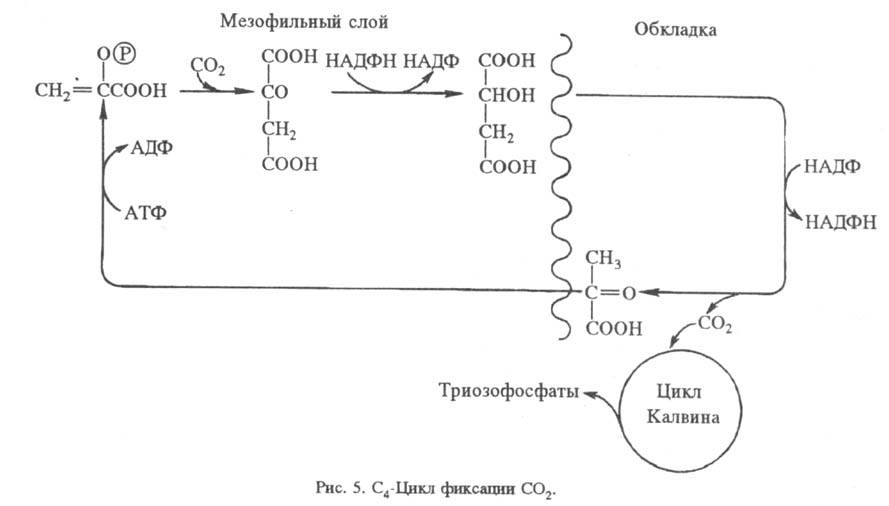
Для кактусов, молочая и др. засухоустойчивых растений характерно частичное разделение фиксации CO2 и Ф. во времени (САМ-обмен, или обмен по типу толстянковых; САМ сокр. от англ. Crassulaceae acid metabolism). Днем устьица (каналы, через которые осуществляется газообмен с атмосферой) закрываются, чтобы уменьшить испарение воды. При этом поступление CO2 также затруднено. Ночью устьица открываются, происходит фиксация CO2 в виде фосфоенол-пировиноградной кислоты с образованием C4-кислот, которые днем декарбоксилируются, а освобождаемый при этом CO2 включается в цикл Калвина (рис. 6).
Ф. галобактерий. Единственный известный в природе не-хлорофилльный способ запасания энергии света осуществляют бактерии Halobacterium halobium. Ha ярком свету при пониженной концентрации O2 они образуют в своих мембранах пурпурный белок бактериородопсин. В результате индуцированной светом цис-транс-изомеризации ретиналя (хромофора этого пигмента) происходит поглощение H+ и синтез АТФ. Последний используется для частичного обеспечения энергетич. потребностей клетки.
Историческая справка. Ок. 1770 Дж. Пристли обнаружил, что растения выделяют O2. В 1779 Я. Ингенхауз установил, что для этого необходим свет и что O2 выделяют только зеленые части растений. Ж. Сенебье в 1782 показал, что для питания растений требуется CO2; в нач. 19 в. H. Соссюр, исходя из закона сохранения массы, подтвердил, что большая часть массы растений создается из CO2 и воды. В 1817 П. Пельтье и Ж. Каванту выделили зеленый пигмент хлорофилл. Позже К.А. Тимирязев показал близость спектра действия Ф. и спектра поглощения хлорофилла. Ю. Сакс в сер. 19 в., по-видимому, первым осознал, что этот продукт накапливается в хлоропластах, а Т.В. Энгельман доказал, что именно там же выделяется и O2.
В работах Ф. Блэкмана (1905), P. Эмерсона и У. Арнолда (1932), а также P. Хилла (1936–41) показано наличие световой и темновой стадий Ф. и экспериментально реализована световая стадия в отсутствие CO2 с использованием искусств. акцепторов электрона. Тем самым были получены подтверждения представлений об образовании O2 путем окисления воды. Окончательно это было доказано масс-спектрометрич. методом (С. Рубен, M. Камен, а также А.П. Виноградов и Р.В. Тейс, 1941).
В 1935–41 К. Ван Ниль обобщил данные по Ф. высших растений и бактерий и предложил общее уравнение, охватывающее все типы Ф. Х. Гаффрон и К. Воль, а также Л. Дёйсенс в 1936–52 на основе количеств. измерений выхода продуктов Ф. поглощенного света и содержания хлорофилла сформулировали представление о "фотосинтетич. единице" — ансамбле молекул пигмента, осуществляющих светосбор и обслуживающих фотохим. центр.
В 40-50-х гг. M. Калвин, используя изотоп 14C, выявил механизм фиксации CO2. Д. Арнон (1954) открыл фотофосфорилирование (инициируемый светом синтез АТФ из АДФ и H3PO4) и сформулировал концепцию электронного транспорта в мембранах хлоропластов. P. Эмерсон и Ч.M. Льюис (1942–43) обнаружили резкое снижение эффективности фотосинтеза при 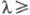 700 нм (красное падение, или первый эффект Эмерсона), а в 1957 Эмерсон наблюдал неаддитивное усиление Ф. при добавлении света низкой интенсивности с
700 нм (красное падение, или первый эффект Эмерсона), а в 1957 Эмерсон наблюдал неаддитивное усиление Ф. при добавлении света низкой интенсивности с 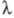 650 нм к дальнему красному свету (эффект усиления, или второй эффект Эмерсона). На этом основании в 60-х гг. сформулировано представление о последовательно действующих фотосистемах в электронтранспортной цепи Ф. с максимумами в спектрах действия вблизи 680 и 700 нм.
650 нм к дальнему красному свету (эффект усиления, или второй эффект Эмерсона). На этом основании в 60-х гг. сформулировано представление о последовательно действующих фотосистемах в электронтранспортной цепи Ф. с максимумами в спектрах действия вблизи 680 и 700 нм.
Осн. закономерности образования O2 при окислении воды в Ф. установлены в работах Б. Кока и П. Жолио (1969–70). Близится к завершению выяснение мол. организации мембранного комплекса, катализирующего этот процесс. В 80-х гг. методом рентгеновского структурного анализа детально изучена структура отдельных компонентов фотосинтетич. аппарата, включая реакционные центры и светособирающие комплексы (И. Дайзенхофер, Х. Михель, P. Хубер).
Лит.: Клейтон Р., Фотосинтеч. Физические механизмы и химические модели, пер. с англ., М., 1984; "Ж. Всес. хим. общества им. Д.И. Менделеева", 1986, т. 31, № 6; Фотосинтез, под ред. Говинджи, пер. с англ., т. 1–2, М., 1987; Итоги науки и техники, сер. Биофизика, т. 20–22, М., 1987.
М. Г. Гольдфельд
Значения в других словарях
- Фотосинтез — (от Фото... и Синтез образование высшими растениями, водорослями, фотосинтезирующими бактериями сложных органических веществ, необходимых для жизнедеятельности как самих растений, так и всех др. Большая советская энциклопедия
- фотосинтез — Образование клетками высших растений, водорослей и некоторыми бактериями органического вещества при участии энергии света. Происходит с помощью пигментов (хлорофиллов, бактериохлорофиллов и каротиноидов)... Микробиология. Словарь терминов
- фотосинтез — -а, м. бот. Образование в клетках зеленых растений, водорослей и в некоторых микроорганизмах углеводов из углекислоты и воды под действием света, поглощаемого светочувствительным пигментом, главным образом хлорофиллом (сопровождается выделением свободного кислорода). Малый академический словарь
- фотосинтез — Способность фототрофных бактерий к фотосинтезу, как и у растений, определяется наличием магнийсодержащих порфириновых пигментов — хлорофиллов. Состав бактериальных хлорофиллов, называемых бактериохлорофиллами, отличается от хлорофиллов растений (рис. Биологическая энциклопедия
- фотосинтез — орф. фотосинтез, -а Орфографический словарь Лопатина
- фотосинтез — Фото/си́нтез/. Морфемно-орфографический словарь
- фотосинтез — ФОТОСИНТЕЗ (от греч. phоs, род. падеж photos — свет ц synthesis — соединение), синтез органич. вещества растениями (т. н. фотосинтетиками), в процессе которого происходит преобразование энергии солнечного света в энергию химич. связей. Ветеринарный энциклопедический словарь
- фотосинтез — Фотосинтез — процесс синтеза органических веществ из углекислоты и воды за счет световой энергии. Свойствен высшим растениям, водорослям и бактериям фотосинтезирующим (см. фотосинтез бактериальный). Толковый словарь по почвоведению
- фотосинтез — ФОТОСИНТЕЗ (от греч. phos, род. падеж photos — свет и synthesis — соединение, составление), образование зелёными р-ниями и фотосинтезирующими бактериями необходимых для жизни органич. в-в за счёт энергии Солнца; осн. Сельскохозяйственный словарь
- фотосинтез — ФОТОСИНТЕЗ — окислительно-восстановительная реакция синтеза органических веществ из неорганических с помощью световой энергии, улавливаемой хлорофиллом. Ботаника. Словарь терминов
- ФОТОСИНТЕЗ — ФОТОСИНТЕЗ, химический процесс, возникающий в зеленых растениях, водорослях и многих бактериях, при котором вода и УГЛЕКИСЛЫЙ ГАЗ превращаются в КИСЛОРОД и продукты питания растений при помощи энергии, поглощаемой из солнечного света. Научно-технический словарь
- фотосинтез — Образование клетками растений органического вещества из углекислоты и воды при участии энергии света. Характерная особенность клетки зелёного растения – присутствие пластид – мелких белково-липидных телец, несколько более плотных... Биология. Современная энциклопедия
- фотосинтез — (от фото... и греч. synthesis — соединение), образование клетками высших растений, водорослей и нек-рыми бактериями органич. веществ при участии энергии света. Биологический энциклопедический словарь
- Фотосинтез — Химический процесс, идущий в зеленых растениях под действием световой энергии с помощью пигментов (хлорофиллов и др.). При этом из углекислого газа и воды образуется глюкоза и выделяется кислород. Экологические термины и определения
- фотосинтез — ФОТОСИНТЕЗ, а, м. (спец.). У растений и нек-рых микроорганизмов: биологический процесс превращения лучистой энергии Солнца в органическую (химическую) энергию. | прил. фотосинтетический, ая, ое. Толковый словарь Ожегова
- фотосинтез — ФОТОС’ИНТЕЗ, фотосинтеза, мн. нет, ·муж. (бот., биол.). Процесс создания при помощи света органических соединений из углекислоты воздуха или воды, происходящий в растениях, которые содержат хлорофилл. см. фото... во 2 ·знач. Толковый словарь Ушакова
- фотосинтез — ФОТОСИНТЕЗ [тэ], -а; м. Спец. Процесс образования углеводов из углекислоты и воды под действием света, поглощаемого хлорофиллом, в клетках зелёных растений, водорослей и некоторых микроорганизмов. ◁ Фотосинтетический, -ая, -ое. Ф-ая деятельность. Толковый словарь Кузнецова
- фотосинтез — [см. фото… + cuнmeз] – процесс создания зелёными растениями, содержащими хлорофил, органических веществ из неорганических (углекислота, вода) при помощи энергии, доставляемой лучами света Большой словарь иностранных слов
- ФОТОСИНТЕЗ — ФОТОСИНТЕЗ (от фото... и синтез) — превращение зелеными растениями и фотосинтезирующими микроорганизмами лучистой энергии Солнца в энергию химических связей органических веществ. Происходит с участием поглощающих свет пигментов (хлорофилл и др.). Большой энциклопедический словарь
- фотосинтез — фотосинтез м. Процесс создания в клетках зелёных растений, водорослей и некоторых микроорганизмов углеводов из углекислоты и воды под воздействием света, поглощаемого светочувствительным пигментом (преимущественно хлорофиллом). Толковый словарь Ефремовой
- Фотосинтез — Зеленые растения способны строить (синтезировать) сложные органические вещества из минеральных, пользуясь для этого в качестве источника энергии светом. Как общее правило, исходными материалами для этого процесса... Энциклопедический словарь Брокгауза и Ефрона
