транспортные рибонуклеиновые кислоты
ТРАНСПОРТНЫЕ РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ (тРНК, трансферные РНК, адапторные РНК)
низкомолекулярные РНК, осуществляющие перенос аминокислотных остатков к матричной РНК (мРНК) при трансляции (синтезе полипептидяой цепи на мРНК-матрице в рибосомах).
Присутствуют в цитоплазме и митохондриях. Цитоплаз-матич. тРНК состоят из одной полирибонуклеотидной цепи, включающей 74–95 нуклеотидных остатков (мол. м. 24–31 тыс.), митохондриальные тРНК немного короче. Для всех молекул тРНК характерно присутствие остатка фосфорной кислоты на 5'-конце (см. нуклеиновые кислоты) нуклеотидной цепи (фосфорилирован 5'-гидроксил), наличие последовательности ССА—ОН (С-остаток цитидина, А-аденозина) на 3'-конце и неск. консервативных нуклеотидных остатков, рассеянных вдоль цепи и занимающих во всех тРНК одни и те же положения.
Отличит. особенность тРНК-присутствие в молекуле минорных нуклеозидов (миноров), общее число разновидностей которых в разных тРНК св. 50, а в одной молекуле тРНК их доля может достигать 25%. Они образуются путем метилирования, гидрирования и др. превращений обычных нуклеотидных звеньев в ходе посттранскрипц. модификации тРНК под действием соответствующих ферментов. Большинство миноров влияет на стабильность пространственной структуры и(или) на адапторную функцию тРНК-способность каждой тРНК узнавать свой кодон (участок мРНК из трех нуклеотидных остатков, кодирующий определенную аминокислоту; см. генетический код). На присоединение к тРНК аминокислот (акцепторную функцию) миноры, как правило, не влияют.
Более половины пуриновых и пиримидиновых оснований тРНК с помощью водородных связей образуют внутрицепо-чечные пары по принципу комплементарности (A-U, G-C, G-U; U и G — соотв. остатки уридина и гуанозина), формируя 4 двухспиральных участка. Эти короткие спирали чередуются с участками неспаренных оснований, в результате чего нуклеотидная цепь образует 3 петли. Таким образом формируется вторичная структура, получившая назв. клеверного листа (см. рис.). В ней выделяют: акцепторную ветвь (стебель, черешок) с универсальной 3'-концевой последовательностью, служащей акцептором (местом прикрепления) остатка аминокислоты; дигидроуридиловую ветвь (шпильку), варьирующую по числу входящих в ее состав нуклеотидов и содержащую до 3 остатков дигидро-уридина (DHU); антикодоновую ветвь (шпильку) с петлей в 7 нуклеотидных остатков, в центре которой находится анти-кодон (тринуклеотид, комплементарный кодону мРНК и обусловливающий специфичность тРНК к этому кодону); тимидилпсевдоуридиловую ветвь, или Ty-шпильку, содержащую минорные нуклеозиды риботимидин и псевдоури-дин.
Кроме того, у всех тРНК между Ty-шпилькой и акцепторным стеблем имеется вариабельная петля (V-петля). Число составляющих ее нуклеотидов у разл. тРНК варьирует от 3 до 20. Если петля длинная, то формируется дополнит. пятый двухспиральный участок тРНК, как, напр., у дрожжевых тРНКSer и тРНКLeu (в верхнем индексе-условные обозначения аминокислот, к которым специфичны данные тРНК; букв. обозначения см. в ст. аминокислоты).
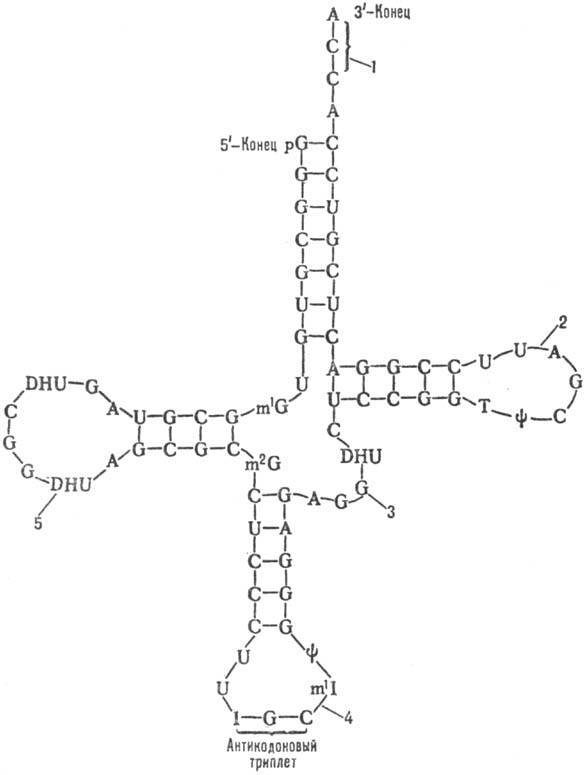
Нуклеотидная последовательность и вторичная структура дрожжевой аланиновой тРНК; линии между антипараллельными участками обозначают водородные связи между комплементарными парами оснований (р-остаток фосфорной кислоты); молекула содержит семь минорных нуклеозидов: y-псевдо-уридин, 1-инозин, Т-риботимидин. DHU-5,6-дигидроуридин, m1I-1-метили-нозин, m1G-1-метилгуанозин, m2G-N2-диметилгуанозин: 1-акцепторная ветвь, 2-Тy-шпилька, S-V-петля, 4-антикодонная ветвь, 5-дигидроуриди-ловая ветвь.
Все тРНК имеют сходную пространственную укладку цепи, напоминающую лат. букву L. Акцепторная и тими-дилпсевдоуридиловая ветви расположены по одной оси, формируя непрерывную двойную спираль, состоящую из 12 пар нуклеотидных остатков; антикодоновая и дигидроури-диловая ветви располагаются также по одной оси, формируя вторую двойную спираль, включающую 9 пар нуклеотидных остатков. Эти два спиральных участка располагаются под углом ок. 90° друг к другу. Трехмерная структура поддерживается нековалентными связями между Ty- и DHU-шпильками, а также др. взаимод., в т. ч. с ионами Mg2+. Конформация тРНК в растворе в целом соответствует ее конформации в кристалле. Важная особенность структуры тРНК заключается в том, что антикодон, находящийся в центре полинуклеотидной цепи и на одном из концов "L", доступен для контактов с мРНК.
В присут. АТФ, ионов Mg2+ и аминоацил-тРНК-синтетаз к группе 3'-ОН 3'-концевого аденозина тРНК присоединяется остаток аминокислоты с образованием аминоацил-тРНК. Аминоацил-тРНК в рибосоме с помощью антико-дона комплементарно связывается с соответствующим ко-доном мРНК. тРНК, акцептирующие разл. аминокислоты, имеют разные последовательности оснований, благодаря чему синтетазы легко их узнают. Через взаимод. кодон-антикодон осуществляется перевод нуклеотидной последовательности мРНК в специфич. аминокислотную Последовательность синтезируемой полипептидной цепи.
Ошибка в узнавании аминокислоты своей тРНК при синтезе аминоацил-тРНК не может быть исправлена на последующих этапах белкового синтеза; последовательность аминокислотных остатков в синтезируемой полипептидной цепи определяется мРНК и аминоацил-тРНК, взаимодействующими в рибосоме, а не природой аминокислотного остатка, связанного с тРНК. Для большинства тРНК с короткой V-петлей важную роль при взаимном узнавании фермента и тРНК играет антикодон, для тРНК с длинной V-петлей-двухспиральные участки. При узнавании происходят взаимные конформац. изменения тРНК и фермента.
Как правило, каждая аминокислота имеет неск. соответствующих ей разновидностей тРНК, незначительно различающихся по первичной структуре и наз. изоакцепторными; их подразделяют на мажорные (доминирующие) и минорные (малочисленные). Структурные различия обусловлены заменами неск. нуклеотидов (или пар нуклео-тидов) в разл. частях молекулы (в т. ч. в антикодоне) и существенно не отражаются на укладке цепей. Для считывания разных кодонов мРНК, соответствующих одной и той же аминокислоте, используются изоакцепторные тРНК с разными антикодонами. Для мн. аминокислот число соответствующих им изоакцепторных тРНК с разными антикодонами гораздо меньше, чем общее число кодонов (напр., 24 митохондриальных тРНК достаточно для узнавания 61–62 смысловых кодонов мРНК). Из этого следует, что одна и та же тРНК может узнавать неск. кодонов, кодирующих одну и ту же аминокислоту, но различающихся по одному нуклеотиду.
Число генов, кодирующих тРНК для одной и той же аминокислоты, может различаться у разных организмов более чем на порядок. Общее число генов тРНК в разл. организмах сильно варьирует (напр., у кишечной палочки Escherichia coli их ок. 70, у шпорцевой лягушки Xenopus laevis ок. 7 тыс., у человека св. 1 тыс.). При транскрипции (синтез РНК на ДНК-матрице) генов тРНК с помощью фермента РНК-полимеразы III образуются предшественники тРНК (пре-тРНК). Дальнейшее их превращение в тРНК включает ряд ферментативных реакций, приводящих к уменьшению размеров молекул и модификации некоторых нуклеозидов. В-генах тРНК эукариот функционально важный 3'-концевой триплет не кодирован-он достраивается посттранскрипционно с помощью фермента тРНК-нуклео-тидилтрансферазы.
Помимо акцепторно-адапторной функции в белковом синтезе, мн. тРНК выполняют роль затравки при обратной транскрипции (синтезе ДНК на РНК-матрице) благодаря комплементарности 3'-конца тРНК (17–20 нуклеотидов) и участка РНК ретровирусов, а также др. ретротранспозонов. На 3'-концах РНК мн. вирусов растений присутствуют тРНК-подобные структуры, обладающие акцепторной активностью. Некоторые тРНК участвуют в биосинтезе пептидогликанов (компонентов внеш. оболочки некоторых бактерий), в переносе аминокислот через внеш. мембрану клеток, в регуляции биосинтеза ряда аминокислот, в посттрансляционной модификации белков (перенос аминокислотного остатка от аминоацил-тРНК на N-конец полипептидной цепи под действием ферментов аминоацил-тРНК-протеин трансфераз), а также во внутриклеточной деградации белков. Имеются данные об участии тРНК как кофактора в реакции восстановления глутаминовой кислоты при биосинтезе хлорофилла. Успехи в изучении структуры и функции тРНК сыграли исключит. роль в понимании общих принципов структурной организации нуклеиновых кислот, в познании биосинтеза белков.
В 1955 Ф. Крик предсказал существование в клетках малых молекул, ковалентно связывающихся с помощью особых ферментов с аминокислотами и участвующих в адаптации (приспособлении) аминокислот к генетич. коду, записанному в нуклеотидной форме (т. наз. адапторная гипотеза). Такими молекулами оказались тРНК и "рН5-фер-менты", названные позднее аминоацил-тРНК-синтетазами. тРНК открыли в 1957 М. Хоглэнд, М. Стефенсон и П. Замеч-ник (США) и одновременно К. Огата и Х. Нохара (Япония).
Впервые нуклеотидную последовательность тРНК установили в 1965 Р. Холли с сотрудниками (США) для дрожжевой тРНКAlа. За последующие 25 лет была расшифрована первичная структура сотен тРНК из разл. организмов (бактерии, дрожжи, млекопитающие и др.). В 1974 А. Рич с сотрудниками (США) и А. Клуг с сотрудниками (Великобритания) впервые с помощью рентгеноструктурного анализа установили трехмерную структуру дрожжевой тРНКРhе в кристалле. Позже трехмерные структуры некоторых др. тРНК были расшифрованы в др. лабораториях.
тРНК-первые нуклеиновые кислоты, для которых была установлена сначала первичная, а затем трехмерная структура их молекул, что имело принципиальное значение для развития мол. биологии, химии прир. соединений и биоорг. химии.
Лит.: Венкстерн Т. В., в кн.: Итоги науки и техники, сер. Молекулярная биология, т. 18, М., 1982, с. 49–109; Киселев Л. Л., Фаворова О. О., Лав-рик О. И., Биосинтез белков от аминокислот до аминоацил-тРНК, М., 1984; Спирин А. С., Молекулярная биология. Структура рибосом и биосинтез белка, М., 1986; Transfer RNA: Structure, properties and recognition, eds. P. Schimmel a. o., N.Y., 1979; Transfer RNA: biological aspects, eds. D. Soil a.o., N.Y., 1980; Schulman L., "Progr. Nucl. Acids, Res. and Mol. Biol.", 1991, v. 41, p. 23.
